




Survival rate of Pacific bluefin tuna (PBT) from hatch to juvenile is less than 2%. One reason of lower survival could be due to limited supply of live feed such as prey fish. To solve this problem, a study was placed to determine suitable formulated commercial diet to replace prey fish showing positive results.
Over the past decade, the high value of bluefin tuna at fish markets in Japan and the rising popularity of healthy foods in Western countries has led to farming and fattening of bluefin tuna in various Mediterranean countries.
Bluefin tuna farming accounted for 18% of global bluefin tuna production (8,954 metric tons) during 2011, and over the same period, tuna accounted for 4% of global seafood production for human consumption. Japan consumes over 75% of global bluefin tuna landings.
Because the production of cultured bluefin tuna mainly depends on wild-captured young fish, an increased exploitation of young fish threatens the bluefin tuna natural resource. Therefore, it is necessary to replace wild-caught young tuna with hatchery-produced juveniles to sustain the tuna farming industry.
“The increase in hatchery production of PBT juveniles has increased the demand for live feed such as yolk-sac larvae of prey fish. The nutritional value of minced sand lance and yolksac larvae is not always stable. Thus, development of formulated diets to replace prey fish and sand lance might increase the production of tuna farming.”
The advantages of formulated diets include freedom of choice in the ingredients and a range of pellet size and nutritional content. An unsuccessful switch in diet could stimulate aggressive behavior and lead to high mortality. It is thus important to determine the effect of switching diets on growth and survival of PBT.
To date, there appears to have been no attempts to report the effect of switching diet on the production performance of PBT larvae. Therefore, it was examined the effect of switching from a single commercial diet plus yolksac larvae during the weaning period, to solely one of three commercial diets after the weaning period, on the survival of PBT larvae.
Materials and Methods
Three commercial diets of marine fish, CM1, CM2, and CM3, were selected as test diets for the feeding trials. These are widely distributed in Japan and used in various marine fish hatcheries. Yolk-sac larvae of the spangled emperor fish L. nebulosus (PF), which is commonly used for juvenile PBT production, were used as the control.
The proximate composition of test diets is shown in Table 1. The crude protein content was approximately 60% in all test diets. Crude lipid content was approximately 18% in all test diets except for CM2, which had slightly lower lipid content (14.5%).

The control diet (PF) contained 66.0% crude protein and 17.5% crude lipid. PBT larvae with an initial size of 20.2 ± 0.3 mm were randomly distributed into eight 500 l circular experiment tanks at a stocking density of 300 individuals per tank (Table 2).

The feeding trial was conducted in duplicate. Fish were sampled at the start of the feeding trial and kept frozen at -80 °C until analysis. At the end of the feeding trial, the remaining fish were stored at -80 °C and transferred to the Laboratory of Fish Nutrition, Tokyo University of Marine Science and Technology, for chemical analysis.
Results
Survival and growth of PBT larvae
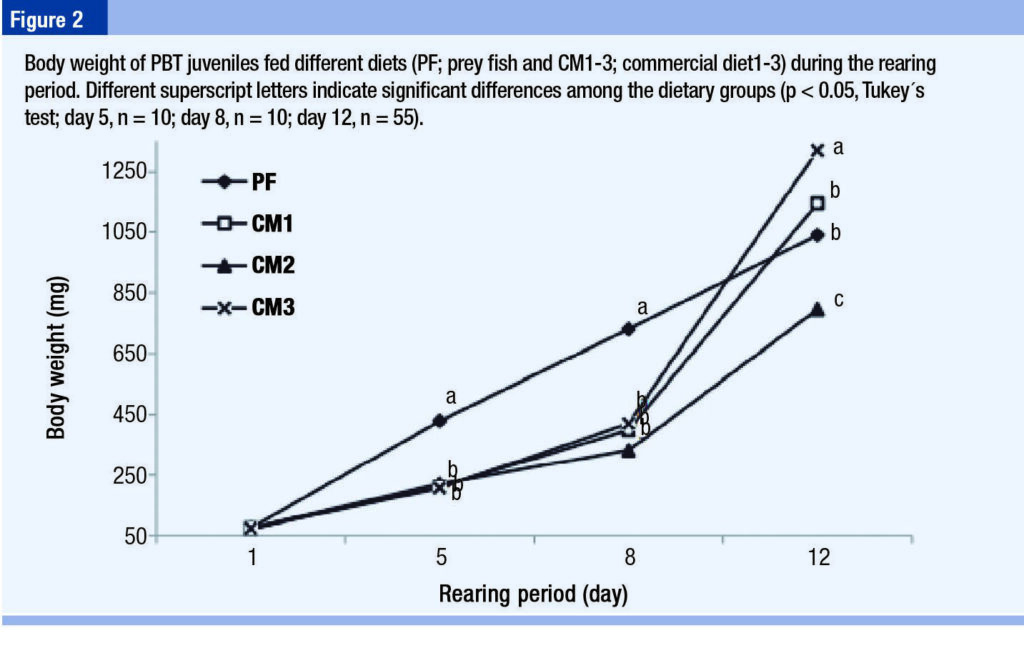
The growth performance of PBT juveniles over the entire rearing period is shown in Figures 1-3. Among the groups, total length and body weight of fish in the CM3 group were significantly greater by the end of the experiment, followed by those in the CM1, PF, and CM2 groups (Figures 1 and 2).

The growth curve for body depth for the PF group was different from the test diet groups (Figure 3). At 32 dph, body depth was significantly greater in the CM3 group than in the PF group (Figure 3).

Proximate chemical composition of the carcass
Among the PF and CM1-CM3 groups, all groups had similar moisture content. No significant difference was observed in protein content among the groups. However, lipid composition was significantly higher in the CM1 and CM3 groups than in the CM2 and PF groups.
“Significant differences among the treatments were observed for arginine, leucine, phenylalanine, alanine, proline, tyrosine, and taurine. High aspartic acid and glutamic acid contents were observed among the examined amino acids.”
The CM3 group had significantly higher arginine, phenylalanine, and leucine contents than the other dietary groups. In PF, the levels of all the amino acids examined, except for glycine and taurine, were as low as nearly one third of those of the other groups.
Although the essential amino acid level of free amino acids in PBT was always higher in the CM1 and CM2 groups than that of the initial fish, histidine, threonine, and valine levels in PBT of the CM3 group were lower than those of the initial fish.
Discussion
Different growth patterns were apparent in the PBT fed test diets and the PF. In the PF group, PBT grew almost linearly throughout the trial in terms of total length and body weight. In apparent contrast, PBT fed test diets showed higher growth rates, particularly between day 8 and 12 of the trial (28-32 dph).
In terms of body depth, the growth rate of the PF group was depressed toward the end of the rearing period. Conversely, as with total length and body weight, growth of PBT fed the test diets was rapid
between day 8 and 12 (28-32 dph).
This difference could reflect the different moisture content between the test diets and PF diet, where a 14.7-fold higher moisture content was observed in the PF diet. This considerable difference might be attributed to he differences in nutrient contents among the PF and test diets, and consequently resulted in higher protein and lipid consumption in the test diet groups.
“Considering that PBT showed rapid growth at approximately 25 dph when fed test diet. Among the groups, by the end of the feeding trial, the highest increase in total length was observed in fish fed CM3.”
Biswas et al. (2009) suggested that a formulated diet with 61.9% crude protein and 17.9% crude lipid could ensure good growth in PBT juveniles. In this experiment, the CM3 diet with 61.1% crude protein and 17.4% crude lipid contained the protein and lipid proportions closest to those recommended by Biswas et al. (2009).
Therefore, the CM3 diet should be feasible as a formulated diet for growing PBT juveniles. The levels of seven amino acids (arginine, leucine, phenylalanine, alanine, proline, tyrosine, and taurine) in PBT were different after being fed the test diet for seven days.
This suggests that these amino acids are affected by the dietary amino acid profile. The CM1and CM3 groups showed higher growth performance than the other dietary groups at 32 dph. The CM1 and CM3 diets contained higher glutamic acid, glycine, and aspartic acid levels than the other diets.
“Higher feeding activity was observed in fish offered CM1, CM3, and PF; these diets had higher alanine, isoleucine, leucine, methionine, and valine levels as free amino acids in relation to CM2.”
The total content of these free amino acids in these diets (0.84 g, 0.68 g, and 0.91 g / 100 g dry-weight for CM1, CM3 and PF, respectively) were higher than that in CM2 (0.19 g / 100 g dry-weight). This difference seemed to affect readily recognition and/or high palatability of the diets and eventually induced higher survival and growth of CM1, CM3 and PF than CM2.
Higher EAA indices and better growth performance was observed in CM1-3 diets. Although highest EAA index was observed in PF group, growth of the PF group was inferior to CM3 group. This could be due to very high moisture content in PF.
PBT showed the best growth in the CM3 group, particularly after 8 days’ post initiation of feeding when growth moved beyond that of the PF group. Considering lower histidine, threonine, and valine among the free amino acids of PBT fed the CM3 diet, these amino acids should be consumed and utilized to support rapid growth after 8 days’ post initiation of feeding.
“The PF group contained the highest DHA and unsaturated fatty acid proportions compared to fish in the test diet groups; this appears to be a reflection of the higher dietary DHA level of PF. Furthermore, the DHA levels of the PBT in the CM1-CM3 groups at 32 dph were lower than that at the start of the experiment at 21dph.”
Conversely, the DHA level at 32 dph of PBT fed PF was higher than that at 21 dph, suggesting that performance of PBT larvae could be affected by difference of DHA. However, similar or even higher growth was observed at 32 dph in PBT fed CM1-CM3 and those fed PF, implying that the DHA level in the test diets was sufficient in terms of growth promotion.
The highest DHA:EPA ratio was observed in the PF diet (6.2), followed by CM3, CM1, and CM2 (2.0, 1.2, and 0.8, respectively). A high DHA:EPA ratio has been reported for marine fish eggs, larvae, and wild copepods that have been considered the natural food of marine fish larvae and juveniles.
Bell et al. (2003) reported that these food organisms have a DHA: EPA ratio > 2, which is closest to the ratios observed in CM3 (2.0) and PF (6.2) in the present study.
“Feeding with CM1 during the weaning period did not appear to negatively affect acceptability of feed, even after the larvae were switched to another diet. In general, it is difficult to evaluate effect of feed on fish performance when fish does not consume the test diet well.”
Therefore, less palatable diet is not suitable for weaning from live feed such as PF to compound diet. In apparent contrast, the result suggests that once PBT were weaned onto CM1, they readily accepted the other diets.
Collectively consideration of the present results therefore suggests that PBT larvae can easily acclimate to a formulated diet once they are successfully weaned onto a different formulated diet with high palatability.
Conclusion
Among the test diet groups, the highest survival occurred for CM1, followed by CM2 and CM3. Of the groups, the CM3 group had significantly greater total length and body weight. These results suggest that PBT larvae and juveniles can readily acclimate to formulated commercial diets once they are successfully weaned onto a suitable formulated diet such as CM1.
This is a summarized version developed by the editorial team of Aquaculture Magazine based on the review article titled “PRODUCTION PERFORMANCE OF PACIFIC BLUEFIN TUNA THUNNUS ORIENTALIS LARVAE AND JUVENILES FED COMMERCIAL DIETS AND EFFECTS OF SWITCHING DIETS” developed by: JEONGHYEON CHO, YUTAKA HAGA – Tokyo University of Marine Science and Technology; YUJI KAMIMURA, ATSUSHI AKAZAWA – Amami Fish Farm Co; AKIRA ITOH – Maruha Nichiro Co and SHUICHI SATOH- Tokyo University of Marine Science and Technology.
The original article, including tables and figures, was published on OCTOBER, 2016, through AQUACULTURE SCIENCE. The full version can be accessed online through this link: https://doi.org/10.11233/aquaculturesci.64.359



