




The vibrios carrying the PirA and PirB toxin genes cause acute hepatopancreatic necrosis disease (AHPND) in cultivated shrimp, significantly affecting the yield. The use of probiotic strains is a promising strategy to prevent vibriosis in shrimp cultures. Here we presented results indicating that the preventive use of probiotic strains with colonization capacity increase larvae protection. This may be a good strategy for the control of AHPND in shrimp larvae hatcheries.
Bacterial diseases produced or induced by species within the Vibrio genus have caused significant economic losses in shrimp farming worldwide due to mass mortality rates up to 100%. A recent example is the acute hepatopancreatic necrosis disease (AHPND) that mainly affects postlarvae and juvenile penaeid shrimp.
Several research’s reveals that AHPND is caused by virulent strains of Vibrio parahaemolyticus carrying the virulent plasmid pVA1 containing PirA and PirB toxin genes. Recently, it has been reported that the bacterial etiology of AHPND also involves other vibrios, including, Vibrio harveryi, Vibrio owensii, Vibrio campbellii, and Vibrio punensis.
“Lai et al. (2015) determined that when the pathogenic AHPND-causing V. parahaemolyticus colonizes the shrimp stomach, it starts to produce Pir A and Pir B toxins. These toxins reach the hepatopancreas, provoking severe injuries.”
In Central America, losses provoked by AHPND has been reported in shrimp larvae hatcheries. In 2017, epidemiological studies performed in South America indicated that strong mortalities registered in hatcheries were associated with V. parahaemolyticus, PirA and PirB toxin carrier.
Vibriosis has traditionally been treated with antibiotics for prophylactic and therapeutic purposes with poor results. Furthermore, antibiotics induce bacterial resistance, even in V. parahaemolyticus, and the risk of the accumulation of residues in the environment.
“In Ecuador, several studies with locally isolated marine bacteria have proven to cause beneficial effects on P. vannamei cultures. In addition, the survival and yield of ponds stocked with shrimp larvae treated with Ili was higher than the ponds stocked with non-treated larvae.”
The Vibrio hepatarius (P62) and the probiotic strain P64, recently identified as Bacillus cereus sensu stricto probiotic strains increased the immune response of P. vannamei juveniles, resulting in a positive effect on survival in treated ponds (Gullian et al., 2004; Aguayo et al., 2009).
Despite the obvious benefits of the probiotic consortium (Ili, P62, and P64), the structures necessary to colonize shrimp surfaces, and the competition for colonization with AHPND-causing V. parahaemolyticus, has not been studied until now.
To fill this gap, the presence of cell surface appendages of bacteria in the probiotic strains was characterized by scanning electron microscopy in transmission mode. Afterwards, the colonization ability of the labeled probiotics in the postlarvae (PLs) of P. vannamei was evaluated using fluorescence and confocal microscopy.
Material and methods
The probiotics V. diabolicus (Ili strain), V. hepatarius (P62 strain), B. cereus s.s. (P64 strain) and the pathogen AHPND-causing V. parahaemolyticus (BA94C2 strain) in shrimp, were provided by the Department of Microbiology of the National Center for Aquaculture and Marine Research (CENAIM, ESPOL).
Initially, the three probiotic strains were characterized by scanning electron microscopy. Next, the colonization capacity of the three probiotic strains in P. vannamei shrimp larvae was determined by direct exposure of the shrimp larvae to probiotics previously stained with fluorochrome dyes, orange acridine, Evans blue, and DAPI (4′, 6-diamidino-2-phenylindole).
“The larvae were observed using epifluorescence and confocal microscopy. Larvae exposed to AHPND-causing V. parahaemolyticus stained with acridine orange were quickly and completely colonized. Finally, the probiotic consortium in P. vannamei larvae was evaluated in a larviculture (Nauplii 5 to postlarvae 8), where the three probiotics were applied daily.”
The trial included a control group without the application of the probiotic. After 16 days of culture, the larvae treated with the microbial consortium and the control group were challenged with a pathogenic, the AHPND-causing V. parahaemolyticus strain.
Results
Morphological characterization of probiotic strains
Scanning electron microscopy microphotographs (Figure 1) focused on the observance of fimbriae and flagella in the cells. Cellularly, Ili is oval, presenting rounded ends with size between 1.3 and 2.2 μm in length and 0.6–0.9 nm in width, with a distinctive presence of numerous lateral flagella (Figure 1A).

P62 has ovalshaped cells with a size between 1.4 and 2.7 μm in length and 0.8 to 1.7 μm in width with a very well-defined plasma membrane structure, where the fimbriae are anchored (Figure 1B). P64 has a characteristic bacillar shape with a size between 1.5 and 2.9 μm long and 0.8–1.2 μm wide, in addition to a well-defined plasma membrane with the presence of flagella and fimbriae (Figure 1C).
All three probiotics were able to form biofilms after 48 h of incubation. However, the strongest biofilm was formed by the probiotic bacteria Ili (p < 0.05) followed by P62 and P64. These results were confirmed when the biofilms were stained with acridine orange (A0).
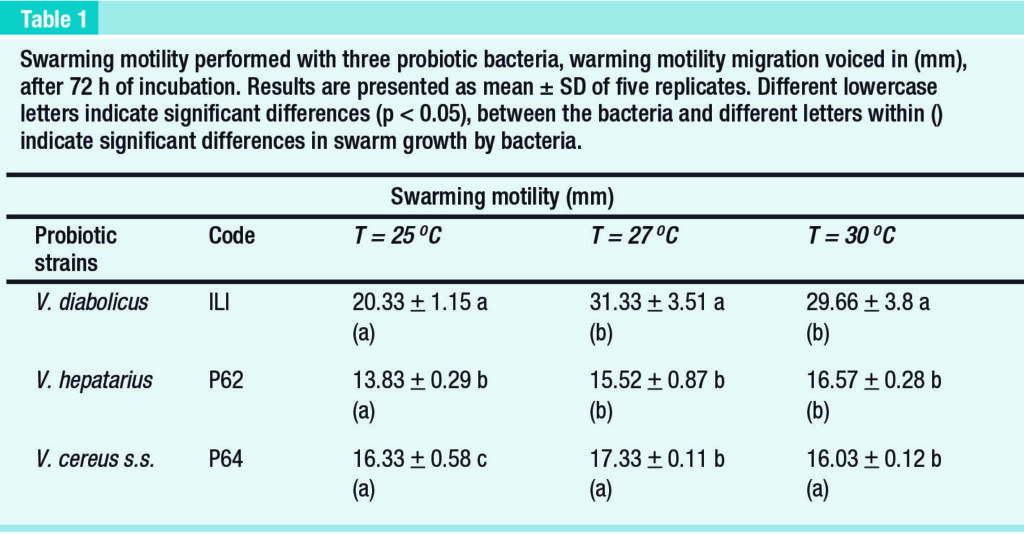
Swarm motility assays of probiotic bacteria
The probiotic Ili and P62 exhibited lower swarming motility at 25ºC. Ili reached its maximum growth at temperatures of 27ºC (p < 0.05). The migration of Ili was significantly higher than of the other probiotic bacteria (p < 0.05) at all temperatures.
The swarming motility of P64 was not affected negatively by low temperature. In addition, at 25ºC the swarming motility of P64 was significantly higher (p < 0.05) as compared to probiotic P62 (Table 1).
Probiotic bacteria colonization assay in shrimp larvae
The confocal microscopy allowed to verify the adhesion capacity of the three probiotic bacteria to the shrimp cuticle and gut, and Figure 2A shows the Ili probiotic strain attached to the oral zone. In Figure 2B, the P62 probiotic strain appear adhered to the peritrophic membrane of intestine.

In the Figure 2C, the probiotic P64 stained with DAPI is detected in the cells of uropod’s. The protecting features of the three probiotics were evaluated by their interaction with AHPND-causing V. parahaemolyticus.
Larvae in Z3 stage exposed to V. parahaemolyticus, were completely colonized, observing larvae strongly stained with AO (Figure 3A). If the larvae were previously colonized with each individual probiotic, the signal of V. parahaemolyticus was less, and it was restricted to the external cuticle and a section of the lateral midgut cecum (Figure 3B, C, D).

When the larvae were colonized by the probiotic consortium, only a weak signal of the pathogenic Vibrio was detected in the external cuticle (Figure 3E and F), and no signal of AHPND-causing V. parahaemolyticus was detected in the digestive system.
The larvae of the negative control exhibited a weak signal of fluorescence, when they were treated with the supernatants of the bacteria stained with AO. Only natural fluorescence was detected in larvae treated with the supernatant (bacteria-free) of the bacteria stained with DAPI and Evans blue.
“In vivo trials, challenge test of shrimp larvae cultivated with the probiotic consortium exposure At the end of the larviculture, the survival rate of the PL8 treated with probiotics consortium was 83.8%, with no significant differences compared to the non-treated larvae (84.4%).”
However, after the challenge test with AHPND-causing V. parahaemolyticus, the survival rate of the larvae treated with the probiotic consortium was 75.33 ± 12.62%, significantly higher (p < 0.05) than the survival rate of the control larvae (32.27 ± 9.57%). Discussion In larviculture environments, pathogenic shrimp vibrios exhibit the ability to proliferate planktonically in the water, as well as rapidly colonize shrimp.
Therefore, probiotic bacteria must be able to compete for these spaces, exhibiting colonization abilities. The microscopic observations, as well as the in vitro and in vivo results achieved in the study, indicate that the probiotics Ili, P62 and P64 possess the cell surface appendages, necessary for the colonization of the host surfaces.
Larvae exposed to probiotic consortium exhibited a high survival after a challenge with AHPND-causing V. parahaemolyticus. Thus, the application of the probiotic consortium in the P. vannamei in hatcheries could provide the larvae with a protective effect against pathogenic Vibrio spp.

Electron microscopy allowed to determine that the robust swarm motility observed in the probiotic Ili could be explained with the large number of lateral flagella that this bacterium has. It is established that the bacterial behavior of swarming motility on solid surfaces is mediated by lateral flagella.
The probiotic P64 is also characterized by having lateral flagella and its swarming motility is greater than P62, in which these cellular appendages were not detected. Lateral flagella not only facilitate swarming motility, but they also contribute to surface adhesion, thereby facilitating the formation of biofilms (Merino et al., 2006).
“This could also explain why more developed biofilms were observed in Ili strain. Another non-flagellar bacterial cell structure that facilitates biofilm formation is the fimbriae. These were very abundant in P62, a bacterium that exhibited more robust biofilms than P64, in which fewer fimbriae were observed.”
Together the three probiotics colonized both the external and internal surfaces of the larvae. The occupation of these sites could be one of the main action mechanisms of these probiotics. In this regard, it should be noted that AHPND-causing V. parahaemolyticus infects shrimp by first colonizing the surfaces of the stomach, where it generates toxins that can penetrate the hepatopancreas, therefore destroying it.
If these sites are already occupied by probiotics, the pathogenicity of V. parahaemolyticus could be reduced. Complete exclusion of AHPND causing V. parahaemolyticus in the gut was obtained when the probiotic consortium was employed. In other words, there was an efficient colonization of the gut and external cuticle by Ili and P62, and colonization of gut and peritrophic membrane of the intestine by P64.
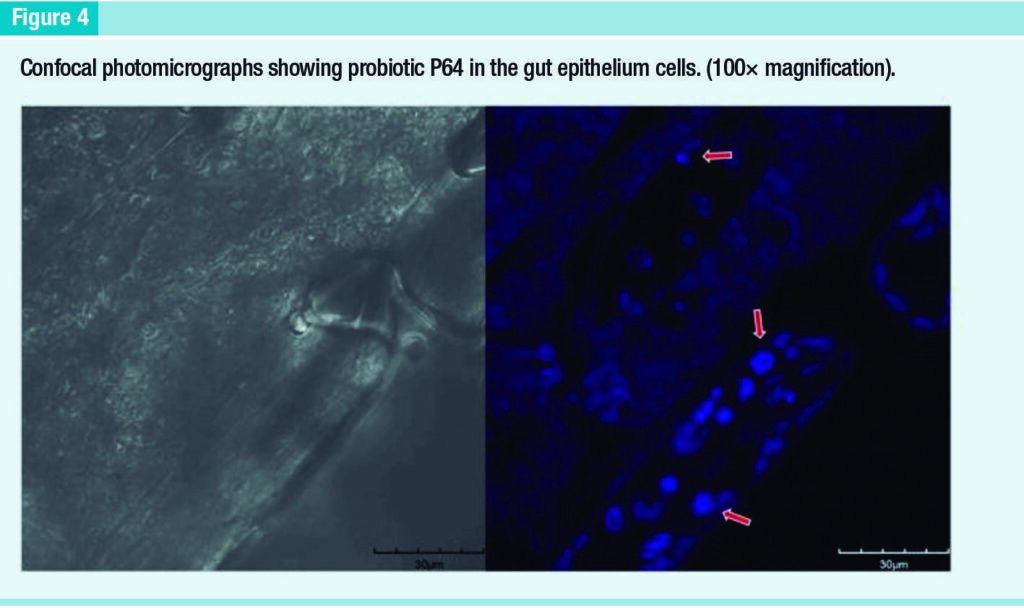
The three probiotics generated a strong fluorescent signal in the lateral midgut caecum (which will differentiate into hepatopancreas). It was detected a strong fluorescent signal for P64 in the base of the uropod setae and in the underlying cells, indicating that this could be a possible route of entry at least for this probiotic (Figure 2C).
In fact, it was observed a strong signal for P64 in the gut epithelium cells (Figure 4). Therefore, it is assuming that these bacteria can penetrate the host’s hemocele.
Shrimp larvae cultivated with the probiotic consortium exhibited an improved survival (75%) when they were exposed to AHPND-causing V. parahaemolyticus. Probiotics attached to external cuticles and internal surfaces with competitive exclusion capacity against pathogenic V. parahaemolyticus may explain these levels of survival.
Conclusion
The probiotics Ili, P62, and P64 possess fimbriae and flagella, cell surface appendages, necessary for the colonization of surfaces through biofilms and swarming motility. These probiotics colonize shrimp larvae excluding the pathogenic AHPND-causing V. parahaemolyticus. These findings are critical to develop new probiotics for the aquaculture industry.
This is a summarized version developed by the editorial team of Aquaculture Magazine based on the review article titled “THE PROBIOTICS VIBRIO DIABOLICUS (ILI), VIBRIO HEPATARIUS (P62), AND BACILLUS CEREUS SENSU STRICTO (P64) COLONIZE INTERNAL AND EXTERNAL SURFACES OF PENAEUS VANNAMEI SHRIMP LARVAE AND PROTECT IT AGAINST VIBRIO PARAHAEMOLYTICUS)” developed by MERY RAMIREZ – Escuela Superior Politécnica del Litoral, ESPOL and Universidad Estatal Península de Santa Elena UPSE; CRISTOBAL DOMÍNGUEZ-BORBOR – Escuela Superior Politécnica del Litoral, ESPOL; , LIZETH SALAZAR; ALEXIS DEBUT; KARLA VIZUETE- Universidad de las Fuerzas Armadas ESPE; STANISLAUS SONNENHOLZNER – Escuela Superior Politécnica del Litoral, ESPOL; FRANK ALEXIS – Universidad Yachay Tech, School of Biological Sciences and Engineering; JENNY RODRÍGUEZ – Escuela Superior Politécnica del Litoral, ESPOL.
The original article, including tables and figures, was published on DECEMBER, 2021, through AQUACULTURE.
The full version can be accessed online through this link: https://doi.org/10.1016/j.aquaculture.2021.737826



