




The use of probiotics, prebiotics, and dietary fiber has become a common practice in aquaculture as an alternative to antibiotic treatment. However, not much is known about the metabolic mechanisms underlying the effects of these products. An evaluation of immune stimulatory products for whiteleg shrimp using a metabolomics approach has shown that the combination of cellulose fiber and probiotics could potentially improve the health and growth of farmed shrimp.
Penaeid shrimp include commercially important species for aquaculture in Asia and the Americas. The whiteleg shrimp, Penaeus vannamei, is the most economically valuable species and its production reached 4.45 tons, valued at USD 26.74 billion in 2017.
Along with the exponential increase in shrimp production, infectious disease outbreaks are major constraints faced by shrimp aquaculture. Most threats to shrimp production worldwide are due to viral pathogens and bacterial pathogens.
“A range of antibiotic applications and other chemical treatments have resulted in mixed reviews and further concerns for associated negative impacts. Thus, more sustainable approaches to mitigate these health issues requires new tools in feed formulation that might better prepare the host’s immune system against pathogens.”
To this end, the addition of various additives including prebiotics, probiotics and symbiotics has recently attracted interest in shrimp aquaculture.

Despite the significant progress in probiotic, prebiotic and symbiotic studies in shrimp aquaculture, the molecular processes underlying the mechanisms of their efficacy remain unclear and need further investigations.
To this end, metabolomics, the study of small molecules (metabolites), is an innovative tool of great applicability to elucidate these complex dynamics.
“The application of metabolomics in aquaculture has just emerged within the last decade and is being applied in fields such as immunology and disease, environmental stress and eco-toxicology, post-harvesting and diet optimization.”
No gas chromatography – mass spectrometry (GC- MS)-based metabolomics studies have been reported for probiotic, prebiotic (or cellulose fibers) and symbiotic investigations in shrimp farming.
In this study, it is applied a GC- MS-based metabolomics approach to compare metabolic responses in hemolymph and gills of whiteleg shrimp (P. vannamei) exposed to different immune stimulatory products, including a cellulose fiber, Vibrio alginolyticus as a probiotics and the combination of the cellulose fiber and probiotics.
It is envisaged that findings herein will contribute to the development of efficient immune stimulation treatments in shrimp aquaculture.
Materials and methods
Experimental design
The experiment was carried out at the Centro Nacional de Acuicultura e Investigaciones Marinas (CENAIM), Escuela Superior Politécnica del Litoral (ESPOL) in San Pedro, Ecuador.
Whiteleg shrimp (P. vannamei) (5.7 ± 0.6 g) were obtained from CENAIM’s Experimental Station (Santa Elena Province, Ecuador) and were acclimatized in the lab for five days.
After acclimation, 10 animals were transferred into each of 40 L tanks assigned as control, cellulose fiber, probiotic and mixture of plant fiber and probiotic (ProFib) treatments (Table 1).

For shrimp in the cellulose fiber treatment, the commercial feed (Skretting, 35% protein) was supplemented with a commercial cellulose fiber (Sigmacell® Cellulose, SigmaAldrich, USA) at a concentration of 100 mg· kg-1.
The V. alginolyticus (ILI) probiotics was administered by immersion in the water to reach a final concentration of 105 UCF· mL-1 in the PB and FB treatments. In the third treatment (ProFib), shrimp were cultured with a combination of plant fiber supplemented feed and probiotic enriched water as described above.
Animals in the control tank were only fed with commercial feed without the addition of cellulose fiber or probiotics.
All animals across control and treatments were fed at 5% biomass. The probiotic bacterial strain V. alginolyticus (ILI) was used in this experiment for the probiotic treatment.
For this purpose, the ILI strain was activated in Petri dishes with Trypticase soy agar and 2% ClNa (TSA + 2% ClNa). The plates were incubated at 28ºC for 24 h. Individual colonies were transferred to LB Broth (ratio 4–5 colonies per 100 mL), and incubated at 28ºC with constant movement (110 rpm) for 12 h.
After the incubation period, the bacterial culture was centrifuged at 4000 g for 10 min at 4ºC, and resulting microbial pellets were resuspended in seawater and stored at room temperature (25ºC). The final concentration (CFU· mL-1) and bacterial viability were determined by plating the bacterial suspension in marine agar.
Finally, the microbial pellet was resuspended in sterilized seawater and stored at room temperature for later use.
Sampling
The experimental conditions were maintained for 5 days, after which 9 shrimp from each treatment and control groups were sampled for metabolomics analysis. Firstly, 200 μL of hemolymph were collected from each animal and placed into a 2 mL cyro-vial and immediately quenched in liquid nitrogen.
Then, gill tissues were cut and collected into 2 mL cyro-vials and snap frozen in liquid nitrogen.
Metabolomics analysis
Metabolites from hemolymph (200 μL) and gills tissues (5 mg of dried tissues) were extracted with cold (-20 ºC) methanol-water and derivatized with methylchloroformate (MCF) with minor modifications. D4-alanine (20 μL of 10 mM) was added into each sample before extraction as an internal standard.
Blank samples containing only 20 μL of 10 mM d4 alanine were extracted together with samples for quality control (QC) purposes. Another type of QC sample was amino acid mixtures (20 μL, 20 mM) that were derivatized as the protocol for the samples.
After derivatization, 10 μL from each sample were pooled together to make a pooled QC sample for each tissue.
The derivatized samples were analyzed with a gas chromatograph GC7890B coupled to a quadrupole
mass spectrometer MSD5977A (Agilent Technologies, CA, USA) with a quadrupole mass selective detector (EI) operated at 70 eV. The system was equipped with a ZB-1701 GC capillary column (30 m × 250 μm internal diameter × 0.15 μm film thickness with a 5 m guard column) (Phenomenex, Torrance, CA, USA).
Results
Metabolite profiles of hemolymph and gills
The Automated Mass Spectral Deconvolution and Identification System (AMDIS) analysis yielded 81 targets from 416 components for both hemolymph and gill tissues.
After annotation, there were 84 and 81 metabolites annotated in hemolymph and gill, respectively. These compounds belong to major categories including amino acids, fatty acids, organic acids and others.
Metabolic responses of hemolymph to immune stimulation
The Principal Components Analysis (PCA) score plots showed a clear separation between the combined treatment and the control (Figure 1).
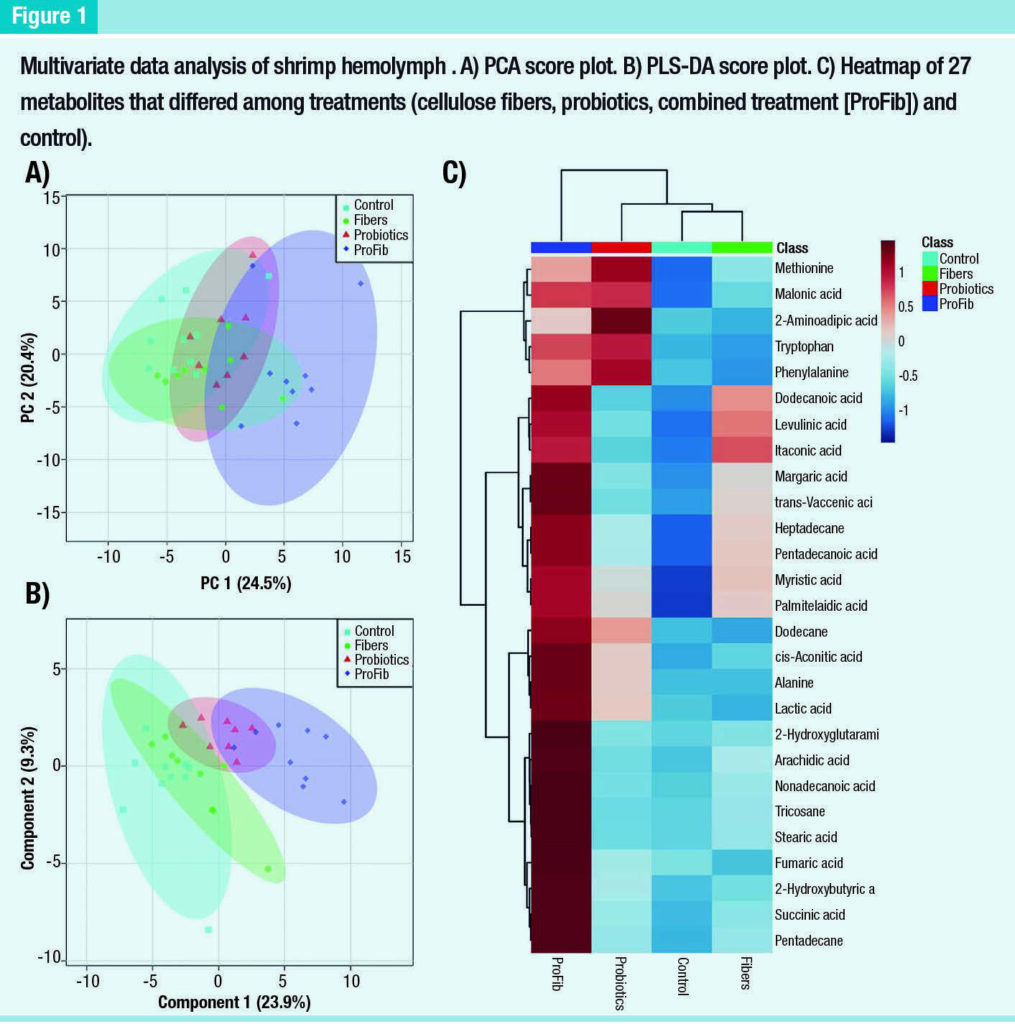
However, there were not clear separations among the control, cellulose fiber and probiotic treatments.
There was some overlap in distribution of the combined treatment, cellulose fiber and probiotic treatment. Similarly, supervised Partial Least Squares – Discriminant Analysis (PLS-DA) did not show a good discrimination between the control and the cellulose fiber treatment, but the separation between the control and the probiotics were clearer than that in the PCA score plot.
The distribution of the combined treatment was clearly discriminated from the control and the cellulose fiber. A one-way ANOVA analysis revealed 27 metabolites that differed among these groups. A heatmap of these metabolites shows the details of these differences (Figure 1), which indicate that most of the differences among groups relate to elevated metabolites in the combined treatment compared to the control.
The probiotics showed higher levels of 5 metabolites (methionine, malonicacid, 2-maninoadipic acid, tryptophan and phenylalanine) and slight increases of 6 other compounds (myrisric acid, palmitelaidic acid, dodecane, cis-aconitic acid, alanine and lactic acid) compared to the control.
Similarly, the cellulose fiber treatment differed from the control with strong increases of 3 metabolites (itaconic acid, levulinic acid and dodecanoic acid) and slight increases of 6 other compounds (margaric acid, trans-vaccenic acid, heptadecane, pentadecanoic acid, myrisric acid and palmitelaidic acid A pathway analysis was performed only between the combined and control groups, since these were the only groups with clear separation in metabolite profiles.
The results revealed 34 pathways involved in this separation. After filtering (impact factor > 0, p ≤ .05, hits ≥ 2), only 6 pathways were found to have significant alterations due to the combined treatment (Figure 2).
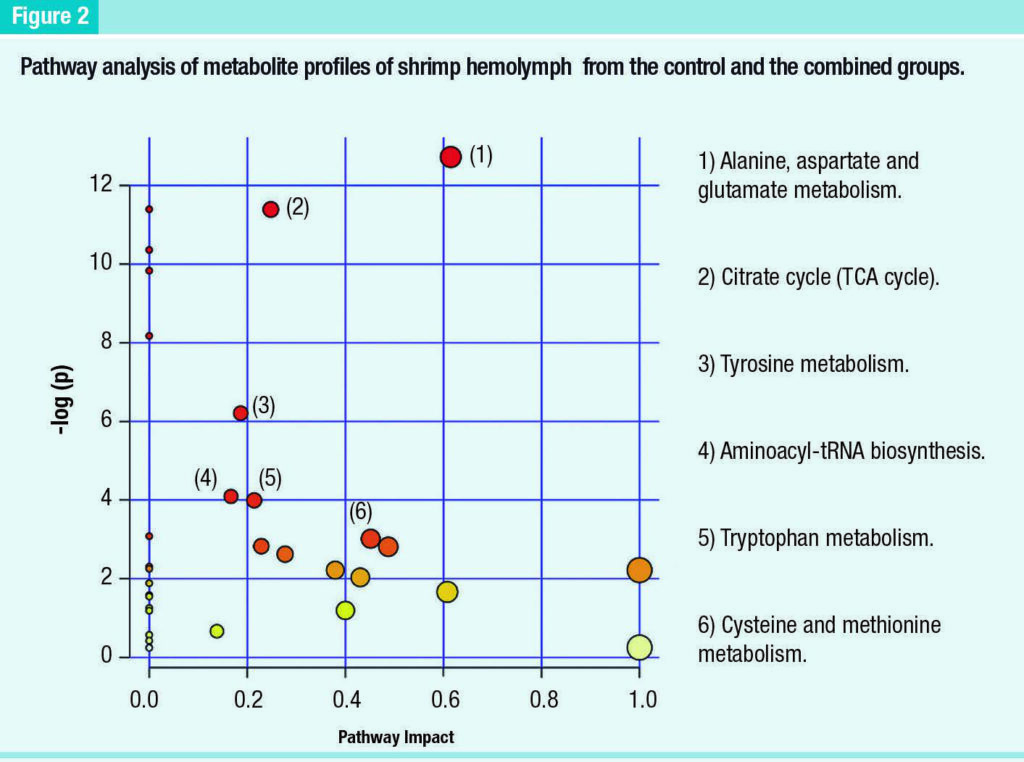
Metabolic responses of gills to immune stimulation
The metabolite profiles of gill tissues did not show clear separation among the treatments via PCA score plot, except for a slight separation with some overlap between the control and the combined group (Figure 3A).
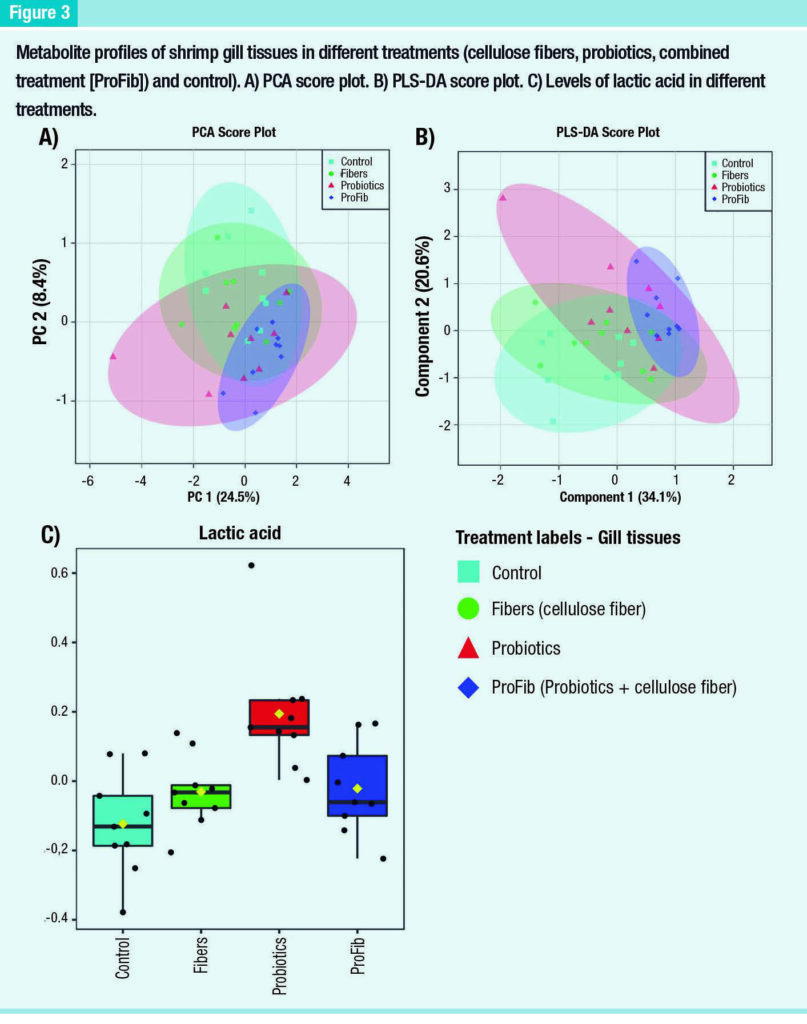
When the PLS-DA analysis was performed, there was a slight separation between the control and combined group, as well as between the control and the probiotic group, but not between the combined and the probiotic groups (Figure 3B).
Similarly, no separation between the cellulose fiber group and the control was observed in the PLS-DA score plot. Consistent with the cluster analysis, a one-way ANOVA revealed only lactic acid as significantly different among the groups (p < 0.05), which was significantly higher in the probiotic treatment than other treatments (Figure 3C).
ROC curve analysis of itaconic acid and lactic acid
Classical univariate ROC curve analyses revealed that the AUC of itaconic acid in hemolymph and lactic acid in both hemolymph and gill tissues had very high values of more than 0.91 (Figure 4). These results suggest that itaconic acid and lactic acid could be important and accurate biomarkers for classification and prediction modeling.

Discussion
This study provides the first GC− MS-based metabolomics investigation on immune stimulation of cellulose fiber (plant fiber), probiotic (V. alginolyticus) and a combined treatment of cellulose fiber and probiotics for whiteleg shrimp (P. vannamei).
Differences in metabolite profiles of shrimp hemolymph and gill tissues revealed significant effects of the cellulose fiber and probiotics on shrimp. While the gill metabolite profiles showed a significant difference only in lactic acid among the treatments, those of hemolymph revealed alterations of 27 metabolites.
“The hemolymph metabolite profiles of individual cellulose fiber and probiotic treatments showed little difference compared to the control, while the combined treatment showed a remarkable difference compared to the control and the other single stimulant treatments.”
This suggests that the best immune stimulation can be achieved with the combined application of the cellulose fiber and probiotics. In agreement with this finding, previous shrimp studies have also shown that the combination of cellulose fiber and probiotic yields higher immune responses and survival of L. vannamei against infections, compared to the individual treatments with either probiotics or prebiotics (Arisa, et.al., 2015 and Huynh, et al., 2018).
Conclusions
This evaluation demonstrates the application of metabolomics to reveal insights into the metabolic responses of shrimp exposed to different immune stimulation products. The combination of cellulose fibers as immunostimulant and V. alginolyticus as probiotics provided a better stimulation than the single stimulants and the control.
The study results suggest that the combination of cellulose fibers and probiotics could potentially
be used in aquaculture to improve the health and growth of farmed shrimp.
“To this end, there is a need for further experiments to investigate the long-term effects of the application of this stimulant on shrimp growth and host responses to pathogen challenges.”
Among the altered metabolites, lactic acid was increased in both hemolymph and gill tissues in shrimp exposed to the combined treatment, but it was the only altered metabolite in the gill tissues of the probiotic treatment group.
This suggests that lactic acid may be a highly sensitive metabolite and it could potentially be used as a stress biomarker for shrimp farm management.
This is a summarized version developed by the editorial team of Aquaculture Magazine based on the review article titled “EVALUATION OF IMMUNE STIMULATORY PRODUCTS FOR WHITELEG SHRIMP (PENAEUS VANNAMEI) BY A METABOLOMICS APPROACH” developed by ANDREA C. ALFARO – Auckland University of Technology, New Zealand; THAO V. NGUYEN – Auckland University of Technology, New Zealand and Nguyen Tat Thanh University, Viet Nam; JENNY A. RODRÍGUEZ, BONNY BAYOT, CRISTOBAL, DOMÍNGUEZ BORBOR and STANISLAUS SONNENHOLZNER – Escuela Superior Politécnica del Litoral, ESPOL, Centro Nacional de Acuicultura e Investigaciones Marinas, CENAIM, Ecuador; AWANIS AZIZAN and LEONIE VENTER – Auckland University of Technology, New Zealand.
The original article was published in FISH AND SHELLFISH IMMUNOLOGY in DECEMBER 2021.
The full version, including tables and figures, can be accessed online through this link: https://doi.org/10.1016/j.fsi.2021.12.007



